Iron Metalloporphyrins Complexes in Bioinorganic Molecules BSc Chemistry Notes
byJitendra Singh Sandhu-
Iron Metalloporphyrins Complexes in Bioinorganic Molecules BSc Chemistry Notes
In these notes we learn about, Iron Metalloporphyrins Complexes in Bioinorganic Molecules. These chemistry notes are helpful for BSc MSc Chemistry students and also helpful for students preparing for CSIR NET, UGC GATE or any other Government exam.
Metalloporphyrins
The metalloporphyrins are bioinorganic molecules in which a metal ion is coordinated to four nitrogen atoms inside the cavity of the porphyrin ring in a square planar geometry.
The axial sites (5th and 6th position) are available for other ligands.
Some examples of metalloporphyrins are hemoglobin, myoglobin, cytochromes and chlorophylls.
The porphyrin rings are the derivatives of a macrocyclic ligand called porphine.
In porphyrin rings various groups are attached to the perimeter of porphine molecule.
The porphyrin rings can accept two hydrogen ions to form the dication (i.e +2 diacid) or donate two protons to form dianion.
In metalloporphyrin complexes the inner hydrogen atoms are replaced as protons by dipositive metal ions.
Therefore, the metal free porphyrin ligand has -2 charges.
Since, this macrocyclic ligand has a planar conjugated system of π-bonds around its perimeter; it is much more rigid macrocyclic ligand than the crown ethers.
Therefore, the ligand is more selective for certain metal atoms than the crown ethers.
It has stronger preferences for the d8 Ni2+ ion.
The other metal ions may add above or below the square plane.
The porphyrin rings are rigid because of the delocalization of the π-electrons around the perimeter.
The size of the cavity in the centre of porphyrin ring is ideal for accommodation of metal ions of the first transition series.
If the metal ion is too small such as Ni2+, the ring becomes ruffled to allow closer approach of nitrogen atoms to the metal ion.
On the other hand, if the metal ion is too large, it cannot fit into the cavity and occupies position above the ring which also becomes domed.
The structures of porphine molecule, metalloporphyrin and Fe-protoporphyrin IX of heme group are shown.
Figure 1Porphine
Figure 2 Metalloporphyrin
Figure 3 Fe-protoporphyrin IX
Role of Iron in body or Role of Iron in living systems
Iron is the most important transition metal involved in living systems, being vital for both plants and animals.
In the living systems, iron has three well characterized systems:
Proteins that contain one or more porphyrin rings such as hemoglobin, myoglobin and cytochrome P-450.
Proteins that contain non- heme iron such as iron-sulphur compounds (ruberdoxin, ferredoxins, and nitrogenase).
The non-heme di-iron oxo-bridged compounds such as carboxylates (hemerythrin, ribonucleotide reductase and methane monooxygenase).
A porphyrin ring containing an Iron (Fe) atom is called a heme group. Each hemoglobin molecule is made up of four subunits, each of which consists of a globin protein in the form of folded helix or spiral. Hemoglobin is called a tetramer because of its four subunits.
The globin proteins are of two types:
Two alpha protein (141 amino acids)
Two beta protein ( 146 amino acids)
Each protein consists of one polar and one non-polar group.
Figure 4: Structure of Haemoglobin
Figure 5: Structure of Haemoglobin
· Made up of four globin protein subunits (a and b)
· Each protein partially encloses a heme group.
· It consist of four subunits;
(a) 2 alpha chains that are each 141 amino acids
(b) 2 beta chains that are each 146 amino acids
· Each heme group lies in a porphyrin pocket.
(a) One axial position of the iron is bound to imidazole nitrogen from the protein.
(b) One axial position is available/vacant or has H2O bound to it.
Deoxyhemoglobin
Figure 6 Structure of Deoxyhemoglobin
The protein is attached to Fe (II) protoporhyrin IX through imidazole nitrogen of histidine residue in such a way that the polar groups of each protein are on the outside of the structure leaving a hydrophobic interior. Therefore, the heme group is held in water resistant protein pocket.
There is a conformational change of the heme group upon coordination of an oxygen molecule which triggers interconvertion of the T (tense) and R (relax) conformations.
In deoxyhemoglobin iron is coordinated to four nitrogen atoms of the planar protoporphyrin IX and the 5th coordination site is occupied by nitrogen atom on imidazole of a proximal histidine of globin protein. The 6th vacant site trans to the imidazole nitrogen is vacant and reserved for oxygen molecules.
In deoxyhemoglobin, iron is present as high spin Fe (II) with one electron occupying the dx2–y2 orbitals (Figure 7) that points directly toward the nitrogen atoms of protoporhyrin IX.
Figure 7 LFSE splitting diagram for a high spin single Fe2+ metal
The presence of this electron increases the size of Fe (II) in these directions by repelling the lone pair of electrons on nitrogen atoms. As a consequence, Fe (II) becomes too large to fit easily within the hole provide by the planar protoporhyrin IX ring.
The Fe (II) ion is, therefore, lies about 40 pm out of the plane in the direction of the histidine group, and the heme group is slightly bent into a domed shape. The iron atom in deoxyhemoglobin has square based pyramidal coordination.
Figure 8 The changes in Heme of Haemoglobin upon oxygenation
The steric interactions between the histidine residues, the associated globin chain and heme group inhibit the free movement of the iron atom into the porphyrin ring.
Oxyhemoglobin
Figure 9 Structure of Oxyhemoglobin
Although oxygen is not a strong ligand, the coordination of the dioxygen molecule trans to the histidine group as a 6th ligand alters the strength of the ligand field and causes the pairing of electrons on iron without affecting the oxidation state of iron. Therefore, Fe (II) becomes low spin and diamagnetic.
In low spin Fe (II), the six d-electrons occupy the dxy, dyz, dzx orbitals (Figure 10). The dx2-y2 orbital is now empty and the previous effects of an electron present in this orbital in repelling the porphyrin nitrogen atom is diminished.
Figure 10 LFSE splitting diagram for a high spin single Fe2+ metal
Therefore, the size of low spin Fe (II) becomes about 17 pm smaller than high spin Fe (II). Thus, the Fe (II) slips into the hole of an approximately planar porphyrin ring. As the iron slips into the hole, the imidazole side chain of histidine also moves toward Fe atom, and the complex has an octahedral geometry.
The four subunits of hemoglobin are linked with each other through salt bridges between the four polypeptide chains. These salt bridges are formed mainly due to electrostatic interaction between the –NH3+ and –COO– groups present on all the four polypeptide chains of hemoglobin.
The protein structures in hemoglobin consist of a peptide backbone with various side chains. These side chains consist of a variety of non-polar (hydrocarbons), cationic (such as –NH3+) and anionic (such as –COO–) groups.
These salt bridges between the polypeptide chains in hemoglobin are now believed to introduce strain in the molecule. Therefore, the deoxy form of hemoglobin is called tense state (T state).
The movement of iron atom and imidazole side chain of histidine toward the porphyrin plane results in breaking of some of the salt bridges. The breaking of these salt bridges reduces the strain in hemoglobin is called relaxed state (R state).
The T form of deoxyhemoglobin discouraged the addition of the first dioxygen molecules.
The bonding of one dioxygen molecule to a subunit of hemoglobin reduces the steric hindrance in the outer subunits (due to breaking of salt bridges) and therefore encourages the bonding of dioxygen molecules to the iron atom of the second subunit which in turn encourages the third as well as fourth subunits.
The binding of dioxygen molecule is the most difficult in the first subunit and the easiest in the last subunit due to conformational change in the protein chain (or polypeptide chain).
Initial addition of a dioxygen molecule to high spin Fe (II) triggers the oxygenation of deoxyhemoglobin. This is called cooperative effect.
The phenomenon where the addition of dioxygen to one heme subunit encourages addition of the dioxygen molecules to other heme subunits is known as cooperative effects.
Conversely, as one oxygen molecule is removed from oxyhemoglobin the reverse conformational changes occur and successively decrease its affinity for oxygen. Therefore, initial removal of oxygen molecule from deoxyhemoglobin triggers the removal of remaining oxygen molecules (cooperative effect).
Formation of hematin
The naked heme, the iron-porhyrin complex without accompanying the polypeptide chains is oxidized to Fe (II) by dioxygen molecule in aqueous solution and is converted immediately into a stable μ-oxo dimer known as hematin. In hematin iron is high spin Fe (III). The hematin is unstable to transport oxygen.
The polypeptide chain can be removed by treatment of HCL/acetone.
The polypeptide chain in hemoglobin and myoglobin prevents oxidation of Fe (II) because:
The hydrocarbon environment round the iron has a low dielectric constant and is hydrophobic and therefore acts as a non-polar and provides non-aqueous environment.
It provides steric hindrance and does not allow the formation of hematin.
The mechanism of the formation of hematin is as follow:
The first step involves the bonding of the oxygen molecule to Fe (II) of the heme group, PFe (II).
Second step involves the coordination of bound oxygen to second heme group forming μ-peroxo complex.
The third step involves the cleavage of the peroxo complex into two ferryl complexes in which iron is present in +4 formal oxidation states.
In the last step, the ferryl complex combines with another heme group resulting in the formation of hematin.
Picket Fence Porphyrins
In nature, O2 reacts with d-block metal ions to form complexes in which the O-O bond is retained (as in superoxo and peroxo species) these products tend to undergo irreversible decomposition involving rapid O-O bond cleavage and formation of water or oxides.
The formation of bridge oxo dimer in hemoglobin occurs because individual heme unit are attached to the bulky protein molecules and the close approach is impossible.
Figure 11 Formation of oxo dimer in hemoglobin
Sterically hindered Fe (II) complexes such as the ‘picket fence’ porphyrin achieve this protection by preventing a second Fe (II) complex from attacking the distal Oxygen atom of the superoxo species to form a bridged peroxo intermediate.
Interaction with the second iron centre can be prevented by using:
Use of steric inhibition (picket fence)
Use of low temperature
Anchor the Fe complex on silica gel
An example of Picket Fence Porphyrins is shown below:
Figure 12 Picket Fence Porphyrins
The four substituent’s in Picket Fence Porphyrins form a cavity, and figure below shows the binding of O2 within this cavity. The axial ligand is 1-methylimidazole which is structurally similar to a His residue. The system clearly resembles the iron environment in haemoglobin.
Figure 13
Myoglobin
Myoglobin (or deoxy-myoglobin) is a protein which has only one heme group per molecule and serves as an oxygen storage molecule in the muscles.
It has molar mass of about 17000
Myoglobin binds dioxygen molecule more strongly than hemoglobin.
The myoglobin molecule is similar to a single subunit of hemoglobin.
Myoglobin is a five coordinate high spin Fe (II) complex with four of the coordination positions occupied by N-atoms of the porphyrin ring.
The fifth position is occupied by an N-atom of an imidazole group of a histidine residue (a globin protein).
The protein consists of 153 amino acids.
The protein restricts access to the Fe (II) by a second heme and reduces the formation of a hematin like Fe (III) dimer.
The results are that the Fe (II) porphyrin complex survives long enough to bind and release dioxygen molecule.
Such five coordinate heme complexes of Fe (II) are always high spin t42g e2g with one electron occupying the dx2–y2 orbital that points directly towards the four porphyrin nitrogen atoms.
The presence of this electron increases the size of Fe (II) in these directions by repelling the lone pair of electrons of the nitrogen atoms.
The size of Fe (II) is 92pm in the square pyramidal arrangement which is considered to be pseudo-octahedral environment with the sixth ligand removed.
The size of Fe (II) is so large that it cannot fit into the hole of the planar porphyrin ring and therefore it lies about 40 pm away from the plane of the ring.
Therefore, high spin Fe (II) porphyrin complexes (in Hb and Mb) involve puckering and twisting of porphyrin ring.
When a dioxygen molecule binds to Fe (II) at sixth coordination site trans to imidazole group of histidine residue, the complex converts to low spin Fe (II) octahedral complex and the electronic configuration changes to t62g (that is the six d-electrons occupy the dxy, dyz and dzx orbitals leading to dx2–y2 and dz2 orbitals empty).
Figure 14 Molecular orbital of Myoglobin molecule
The previous effect of two electrons occupying the dx2–y2 and dz2 orbitals in repelling the N atoms on X, Y and Z axes diminishes.
Therefore, the low spin Fe (II) ion is smaller (75 pm) and slips into the hole in the planar porphyrin ring.
As the Fe (II) ion moves, it pulls the imidazole group of histidine residue.
Therefore, all the nitrogen atoms (including that of proximal histidine) approach more closely to the Fe (II) ion.
Physiology of Hemoglobin and Myoglobin
Hemoglobin has relatively high affinity for dioxygen at high partial pressure of dioxygen whereas myoglobin has relatively high affinity for dioxygen at lower partial pressure of dioxygen.
In vertebrates dioxygen enters the blood in the lungs or gills where the partial pressure of dioxygen is relatively high and hemoglobin is virtually saturated with dioxygen in lungs.
When hemoglobin carries dioxygen to muscles tissues, it experiences the lower partial pressure of dioxygen and its affinity for dioxygen has fallen off rapidly and in this situation affinity of myoglobin for dioxygen is relatively high.
Therefore, in muscles tissues dioxygen is thermodynamically favourable transferred from hemoglobin to myoglobin.
The reactions occurring in lungs and muscles tissues are as follows:
Hb + 4 O2 —–> Hb(O2)4
In lungs
Hb(O2)4 + 4Mb —–>4Mb(O2) + Hb
In muscle tissues
The oxygenation equilibrium for haemoglobin is represented as:
For hemoglobin n=2.8 in the pH range of physiological importance in muscle tissues.
The exact value of n depends on pH of the biological system.
When n exceeds the unity, the attachment of dioxygen to one heme group of hemoglobin increases the binding constant for next dioxygen which in turn increases the binding constant for the next one and so on.
The exponent n=2.8 indicates that attachment of dioxygen to one heme subunit progressively increases its tendency to bind with the subsequent heme subunits of hemoglobin.
Therefore, the exponent 2.8 reveals the cooperative effect of four heme subunits that produces the curve shown.
If the four heme subunits of hemoglobin act independently, they will give curve identical to that of myoglobin.
The cooperative effect favours the attachment of more dioxygen molecules to the heme subunits of hemoglobin.
Conversely, if only one dioxygen molecule is attached to heme a heme subunit of hemoglobin, is dissociates more readily than from a more oxygenated hemoglobin.
This indicates that at low partial pressure of dioxygen hemoglobin is less oxygenated is less oxygenated and tends to release dioxygen whereas at high partial pressure of dioxygen, hemoglobin is oxygenated almost to the same extent as if n=1.
This results in a sigmoidal curve for oxygenation of hemoglobin.
The curves show that myoglobin binds dioxygen more strongly than the first dioxygen of hemoglobin.
However, the fourth binding constant of hemoglobin is larger than that for myoglobin by a factor of about 50.
Oxygen binding curves for myoglobin and haemoglobin
The O2 binding curves for Myoglobin and Hemoglobin are shown below (Figure 15):
Figure 15 Oxygen binding curves for hemoglobin and myoglobin
The curve for Haemoglobin is sigmoid which indicates that uptake and release of successive O2 molecules is cooperative.
At low O2 partial pressure (in capillaries the partial pressure of O2 is about 40 torr), hemoglobin has a low affinity, which enables it to transfer its O2 to myoglobin.
As the pressure increases, haemoglobin has a greater affinity for O2 and as a result hemoglobin can pick up O2 in the lungs (partial pressure of O2 is about 100 torr).
This change in affinity is attributed to there being two conformations.
The tensed state (T) has a low affinity and the relaxed state (R) has a high affinity.
Deoxy-Hemoglobin is in tensed state (T) and fully loaded oxy-hemoglobin is in relaxed state (R).
The binding of the first O2 molecule to the T-state molecule is weak, after the binding of the first O2, there is a decrease in the size of the Fe allows it to move into the plane of the porphyrin ring. This pulls on the proximal histidine ligand towards the porphyrin ring.
The effect being to push the other Fe atoms closer to their respective ring planes and in so doing convert the protein into the R state.
This increases the affinity for O2 to bin to haemoglobin molecule.
Bohr Effect
The cooperative effect is pH dependent. The affinity of hemoglobin for di-oxygen decreases with decrease in pH. This is called Bohr Effect.
Figure 16 Oxygen dissociation curves for haemoglobin at different pH. Bohr effect haemoglobin
A decrease in pH promotes the dissociation of oxygen from haemoglobin (Figure 16).
The CO2 released in muscle tissues is the end product of breakdown of glucose.
CO2 being acidic, decreases the pH in muscles tissues and lowers the pH.
The greater the muscular activity the more will be the release of CO2.
In the muscles tissues, there is low PO2, low pH and high PCO2, therefore, oxygen is released from haemoglobin.
In the muscle tissue CO2 is produced and is transported to the lungs. Most of the CO2 is transported in the form of soluble HCO3– ions.
CO2 enters the erythrocytes (red blood cells) where the enzyme carbonic anhydrase converts it to H2CO3 which dissociates into the HCO3– and H+ ions.
CO2 + H2O ——> H2CO3 ——–> H+ + HCO3–
The formation of HCO3– ions is facilitated by the protein chains of deoxyhemoglobin which acts as a buffer by picking up the accompanying protons.
The HCO3– ions travel in the solution in the serum of the venous blood back to the lungs.
The release of proton from hemoglobin on oxygenation produces H2CO3 from HCO3– ion.
HCO3- + H+ ——–> H2CO3
The carbonic anhydrase converts H2CO3 into CO2 which is exhaled out.
H2CO3 ——–> CO2 + H2O
The oxygenated form of hemoglobin is more acidic than deoxygenated hemoglobin resulting in dissociation to produce protons from the molecule when dioxygen binds to it. You are learning about Iron Metalloporphyrins Complexes in Bioinorganic Molecules.
Methemoglobin
Figure 17 Structure of Methemoglobin
Methemoglobin is a metalloprotein in which the iron in the heme is in the Fe3+ state, not in the Fe2+ of normal hemoglobin.
Figure 18 Methemoglobin is a metalloprotein
The size of Fe3+ ion is so small that it can fit into porphyrin ring of hemoglobin without binding oxygen and therefore, it prevents transfer of dioxygen.
In human blood a trace amount (about 3%) of methemoglobin is normally produced spontaneously.
However, the Fe3+ ion has an increased affinity for dioxygen binding.
The binding of dioxygen to methemoglobin results in an increased affinity of dioxygen to other three heme subunits that still contain Fe2+ ion within the same hemoglobin molecule.
This leads to an overall reduced ability of the red blood cells to release oxygen to muscle tissues.
The NADH– dependent enzyme methemoglobin reductase converts the methemoglobin back to hemoglobin.
A higher level of methemoglobin causes a disease called as methemoglobinemia which is a disorder.
NO2-/ NO3- make oxidation of Fe faster than its reduction and causes intoxication.
During the formation of oxyhemoglobin and dioxygen, one electron is partially transferred from Fe2+ of heme to the bound oxygen forming a ferric superoxide complex anion.
Figure 19 Formation of oxyhemoglobin
Cytochrome
Cytochromes are found in both plants and animals and serves as electron carries.
They contain heme like prosthetic groups. A prosthetic group is a compound required by an enzyme to facilitate a particular reaction; such compounds are also called coenzymes.
The peptide portion of an enzyme that requires a prosthetic group is called an apoenzyme while complete enzyme with prosthetic group is called the haloenzyme.
There are three main types of cytochromes (Figure 20):
Cytochrome a (Cyt-a)
Cytochrome b (Cyt-b)
Cytochrome c (Cyt-c)
Figure 20 Structure of cytochrome a, cytochrome b, cytochrome c
Structure of Cytochrome C
Figure 21 Structure of cytochrome c
Heme group- Polypeptide chain attached and wrapped around, contains variable number of amino acids.
5th coordination site- N of imidazole moiety of histidine.
As in hemoglobin, the Fe (II) ion of heme group is attached to a N-atom of imidazole ring of histidine residue on one side of the porphyrin plane.
The sixth coordination site of Fe (II) is occupied by a tightly bound S-atom from a methionine residue of a protein.
For this reaction, the cytochromes are inert not only to oxygen but also to the poisons such as CO which affect the oxygen carries.
Mechanism of action
Cytochrome plays an important role in the mitochondrial electron transfer chain.
It has the ability to undergo reversible changes between Fe (III) and Fe (II), which allows them to act as one electron transfer centres.
The reduction potentials for cytochromes increase in the order: cyt-b (0.04 V), cyt-c (0.26 V), and cyt-a (0.28 V).
Reduction potentials E° for Cytochrome
Cytochrome a
0.28 V
Cytochrome b
0.04 V
Cytochrome c
0.26 V
Therefore, the order of electron flow is: cytochrome b —> cytochrome c —-> cytochrome a —-> O2.
The differences in reduction potentials for Fe (II)àFe (III) oxidation results from changes in the porphyrin substituent, changes in the protein and in some cases changes in axial ligands.
The electron transfer reactions allow energy from glucose oxidation to be released gradually and to be stored in the form of adenosine triphosphate (ATP) which is used when required by a cell.
In the mitochondrial electron transfer chain, Cytochrome c accepts an electron from cytochrome c1 and then transfers it to cytochrome oxidase.
Cytochrome oxidase catalyses four electrons reduction of dioxygen.
O2 + 4H+ + 4 e– ———> 2H2O
Electron is transferred by tunnelling through one of the exposed edges of the heme unit.
Cytochrome-c oxidase, the terminal member of the cytochrome chain which binds the inner membrane of the mitochondrion contains cytochrome-a, cytochrome-a3 and two Cu (II) ions (CuA and CuB).
Electron transfer involves the cytochrome-a and CuA, electron being transferred from cytochrome-c to CuA and then to cytochrome-a.
Cytochrome-a3 and CuB are five and three coordinated respectively and therefore bind dioxygen.
Cytochrome-a3 and CuB provide the site for O2 binding and it converts O2 to H2O. It also involved in the pumping H+ across the mitochondrial inner membrane.
This indicates that cytochrome chain involves transport not only of electrons but also of protons across the mitochondrial membrane.
The oxidized form of cytochrome-a3 is susceptible to bind CN– that renders cyanide toxic.
The diagram below shows how ATP is made via electron transfer using Cytochrome (Figure 22).
Figure 22 Formation of ATP in the inner mitochondrial membrane
Cytochrome P-450
Figure 23 Structure of Cytochrome P-450
Cytochrome P-450 are a group of cytochromes found in plants, animals and bacteria.
It is named as pigment that absorbs at 450 nm with their CO complexes.
This is due to π-π* transition (blue to red) and this band is called SORET band.
Cytochrome P-450 facilitates the cleavage of O2 and functions as monooxygenase catalyzing the insertion of oxygen atoms into substances.
Oxygenase is enzyme that inserts oxygen into the substrates.
Monooxygenase inserts one oxygen atom and a dioxygenase inserts two oxygen atoms into the substrates.
Of the many possible substrates, the most important are molecules in which C-H bond is converted to C-OH groups.
Some examples are:
Conversion of a hydrocarbon RH to ROH.
Conversion of an alkene to epoxide.
Conversion of an aldehyde to the carboxylic acid.
Cytochrome P-450 enzymes are found in kidney where they oxidize insoluble hydrocarbons to water soluble R-OH or epoxides compounds which then be excreted in the urine.
One oxygen atom is inserted into an organic substrate and one atom is reduced to H2O.
Difference between Hemoglobin and Cytochrome-P450
The active site in a cytochrome P-450 is heme similar to hemoglobin and myoglobin except that:
Fe is present in Fe (III) state and it is low spin octahedral.
One S-atom of cysteine is coordinated to Fe (III) instead of histidine in the proximal position.
Sixth coordination site is occupied be H2O
The cytochrome P-450 enzymes have low spin octahedral Fe (III) active site.
Cytochrome P 450 enzymes have molar mass of about 50000.
The organic substrate enters a hydrophobic pocket of the protein, near the Fe (III) centre expelling the enter molecule from the iron axial coordination site to give a Fe (III) complex.
Step 2
The Fe (III) complex is reduced by another enzymatic system to give high spin Fe (II) complex.
Step 3
The dioxygen molecule binds to Fe (II) centre like hemoglobin and myoglobin followed by one electron transfer from Fe (II) to dioxygen to form a Fe (III)-superoxo complex.
Step 4
Another electron is added to give Fe (III)-peroxo complex.
Step 5
The protonation of Fe(III)-peroxo complex leads to removing one oxide ion as water leaving an oxyferryl complex Fe(V)=O or oxygen double bonded to Fe(IV) with one electron having been oxidized from the π-HOMO of the porphyrin ring leaving it as a radical cation.
Step 6
The organic substrate (R-H) is oxidized to R-OH with concomitant binding of an H20 ligand to the active site of the metalloenzyme which once again contains low spin Fe (III) centre.
The catalytic cycle for the action of cytochrome P-450 enzyme is shown below (Figure 25):

 Figure 1Porphine
Figure 1Porphine
 Figure 2 Metalloporphyrin
Figure 2 Metalloporphyrin
 Figure 3 Fe-protoporphyrin IX
Figure 3 Fe-protoporphyrin IX





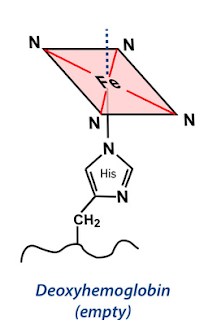 Figure 6 Structure of Deoxyhemoglobin
Figure 6 Structure of Deoxyhemoglobin
 Figure 7 LFSE splitting diagram for a high spin single Fe2+ metal
Figure 7 LFSE splitting diagram for a high spin single Fe2+ metal
 Figure 8 The changes in Heme of Haemoglobin upon oxygenation
The steric interactions between the histidine residues, the associated globin chain and heme group inhibit the free movement of the iron atom into the porphyrin ring.
Figure 8 The changes in Heme of Haemoglobin upon oxygenation
The steric interactions between the histidine residues, the associated globin chain and heme group inhibit the free movement of the iron atom into the porphyrin ring.



 Figure 9 Structure of Oxyhemoglobin
Figure 9 Structure of Oxyhemoglobin
 Figure 10 LFSE splitting diagram for a high spin single Fe2+ metal
Figure 10 LFSE splitting diagram for a high spin single Fe2+ metal






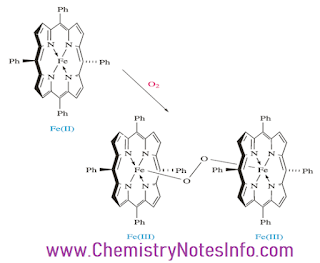 Figure 11 Formation of oxo dimer in hemoglobin
Figure 11 Formation of oxo dimer in hemoglobin
 Figure 12 Picket Fence Porphyrins
Figure 12 Picket Fence Porphyrins
 Figure 13
Figure 13



 Figure 14 Molecular orbital of Myoglobin molecule
Figure 14 Molecular orbital of Myoglobin molecule



 Figure 15 Oxygen binding curves for hemoglobin and myoglobin
Figure 15 Oxygen binding curves for hemoglobin and myoglobin

 Figure 16 Oxygen dissociation curves for haemoglobin at different pH. Bohr effect haemoglobin
Figure 16 Oxygen dissociation curves for haemoglobin at different pH. Bohr effect haemoglobin

 Figure 17 Structure of Methemoglobin
Figure 17 Structure of Methemoglobin
 Figure 18 Methemoglobin is a metalloprotein
Figure 18 Methemoglobin is a metalloprotein
 Figure 19 Formation of oxyhemoglobin
Figure 19 Formation of oxyhemoglobin





 Figure 21 Structure of cytochrome c
Figure 21 Structure of cytochrome c

 Figure 22 Formation of ATP in the inner mitochondrial membrane
Figure 22 Formation of ATP in the inner mitochondrial membrane

 Figure 23 Structure of Cytochrome P-450
Figure 23 Structure of Cytochrome P-450





 Figure 24 Structure of Cytochrome P-450
Figure 24 Structure of Cytochrome P-450

 Figure 25 Catalytic cycle for Cytochrome P-450
Figure 25 Catalytic cycle for Cytochrome P-450

%20(1).png "Interesting Facts About All Elements of Periodic Table - Fun Facts, Fascinating History, and Essential Properties of Every Element")

{kind=link}